scRNA-seq 多样本整合分析教程 (Seurat / Harmony)
环境准备
R 包加载
请选择 common_r 这个环境进行该整合教程的学习
#加载必要的R包
suppressPackageStartupMessages({
library(Seurat)
library(dplyr)
library(ggplot2)
library(patchwork)
library(harmony)
})
# 设置随机种子
set.seed(1234)
# 设置Seurat选项(注意:8000 * 1024^2 实际上是8GB)
options(future.globals.maxSize = 8000 * 1024^2) # 8GBmy36colors <-c( '#E5D2DD', '#53A85F', '#F1BB72', '#F3B1A0', '#D6E7A3', '#57C3F3', '#476D87',
'#E95C59', '#E59CC4', '#AB3282', '#23452F', '#BD956A', '#8C549C', '#585658',
'#9FA3A8', '#E0D4CA', '#5F3D69', '#C5DEBA', '#58A4C3', '#E4C755', '#F7F398',
'#AA9A59', '#E63863', '#E39A35', '#C1E6F3', '#6778AE', '#91D0BE', '#B53E2B',
'#712820', '#DCC1DD', '#CCE0F5', '#CCC9E6', '#625D9E', '#68A180', '#3A6963',
'#968175', "#6495ED", "#FFC1C1",'#f1ac9d','#f06966','#dee2d1','#6abe83','#39BAE8','#B9EDF8','#221a12',
'#b8d00a','#74828F','#96C0CE','#E95D22','#017890')数据读取
本教程提供两种不同形式的数据输入方式,以满足不同用户的数据获取需求,选择适合自己的一种方式即可:
云平台 RDS 文件读取
数据特点:
- RDS 文件是标准的 Seurat 对象文件
- 数据已经过预处理和多个样本合并
- 可直接用于后续的下游分析,也可以提取表达矩阵,重新整合
适用场景:
- 当您无法获得标准的
filtered_feature_bc_matrix表达矩阵时 - 希望整合云平台现有数据进行学习时
- 需要快速重新进行 scRNA-seq 数据整合分析时
注意事项:
- 具体挂载数据和 rds 文件的读取,请参照 jupyter 使用教程
例如下列项目数据/home/demo-SeekGene-com/workspace/data/AY1752565399550/
input <- readRDS("/home/demo-seekgene-com/workspace/data/AY1752565399550/input.rds")
meta <- read.table("/home/demo-seekgene-com/workspace/data/AY1752565399550/meta.tsv", header=TRUE, sep="\t", row.names = 1)
data <- AddMetaData(input,meta)
data <- CreateSeuratObject(counts = input@assay$RNA@counts, meta.data=meta)
seurat_list <- SplitObject(data, split.by = "Sample")
#删除多余的数据,清内存
rm(data,input)
gc()标准 filtered_feature_bc_matrix 文件读取
适用场景:
- 当您拥有标准的基因表达矩阵文件时
- 希望自主完成 scRNA-seq 数据的多样本整合和批次矫正时
- 需要进行完整的从原始数据到整合分析的工作流程时
注意:
- 请保证样本与样本之间的文件结构如下:
scRNA-seq 数据(基因表达矩阵)
├── S127/
│ ├── filtered_feature_bc_matrix/
│ │ ├── barcodes.tsv.gz(细胞条形码)
│ │ ├── features.tsv.gz(peaks 列表)
│ │ └── matrix.mtx.gz(稀疏计数矩阵)
├── S44R/
│ ├── filtered_feature_bc_matrix/
│ │ ├── barcodes.tsv.gz
│ │ ├── features.tsv.gz
│ │ └── matrix.mtx.gz
# 定义样本名称
sample_names <- c('S127', 'S44R')
# 创建一个空的list来存储每个样本的Seurat对象
seurat_list <- list()
for (sample in sample_names) {
# 构建文件路径
data_path <- file.path(sample, 'filtered_feature_bc_matrix')
# 读取表达矩阵文件
counts <- Read10X(data.dir = data_path)
# 创建Seurat对象
rna_obj <- CreateSeuratObject(
counts = counts,
project = sample,
min.cells = 3, # 过滤在少于3个细胞中表达的基因
min.features = 200 # 过滤表达基因少于200个的细胞
)
# 添加样本标识
rna_obj$Sample <- sample
rna_obj$orig.ident <- sample
# 将Seurat对象添加到list中
seurat_list[[sample]] <- rna_obj
# 清除rna_obj对象
rm(rna_obj)
gc()
}质量控制
在整合前,需要先把各样本数据分别"清洗干净":
- QC(质量控制):剔除低质量细胞(如线粒体占比过高、背景或者双细胞等)
质控指标计算
为每个样本计算关键的质控指标,用于后续的细胞过滤。
常用 QC 指标:
nCount_ATAC:总 ATAC 计数,极低或极高都需警惕异常。percent.mt: 线粒体基因比例(通常<20%)nFeature_RNA: 总 ATAC 特征数(通常在 200-10000 之间)nCount_RNA: 总 ATAC 特征数(通常在 200-10000 之间)
# 对seurat_list中的每个对象进行质量控制
seurat_list <- lapply(seurat_list, function(x) {
x[["percent.mt"]] <- PercentageFeatureSet(x, pattern = "^MT-")
return(x)
})质控指标可视化
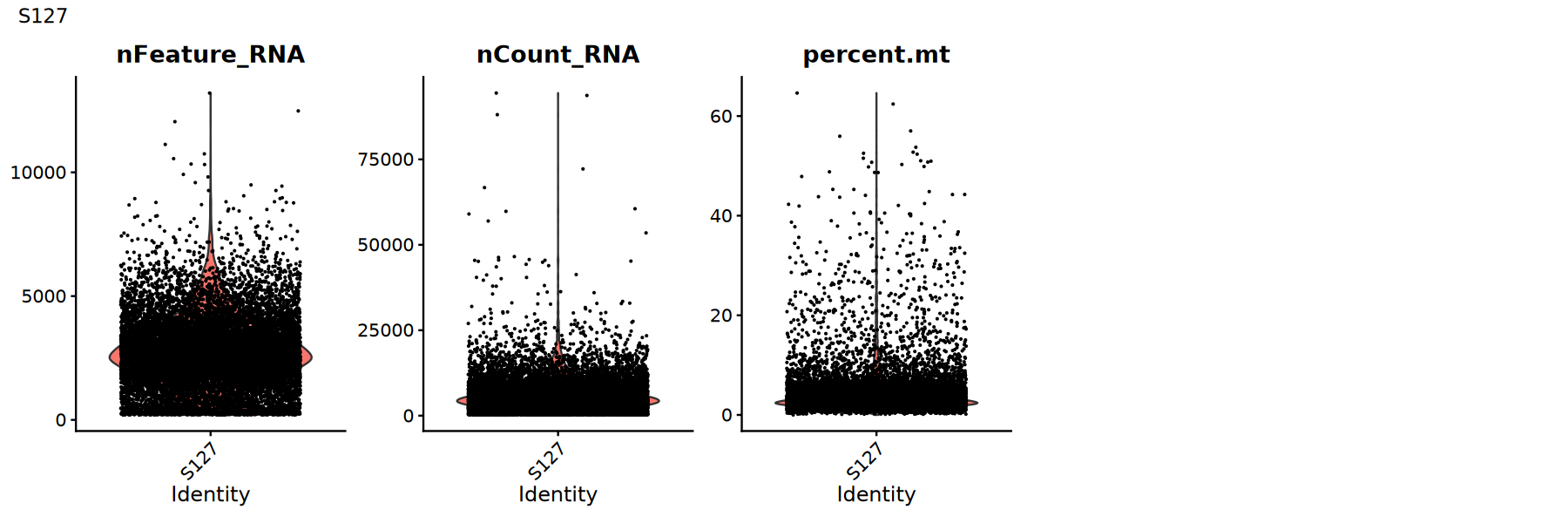
用小提琴图查看各 QC 指标的分布/异常点,确定合适的阈值:
建议:
- 观察是否存在明显的长尾或双峰分布
- 尝试多组阈值并比较下游聚类/UMAP 是否更清晰
options(repr.plot.width = 15, repr.plot.height = 5)
for (sample_name in names(seurat_list)) {
seurat_obj <- seurat_list[[sample_name]]
# 小提琴图展示质控指标分布
p1 <- VlnPlot(
object = seurat_obj,
features = c('nFeature_RNA', 'nCount_RNA', 'percent.mt'),
pt.size = 0.1,
ncol = 5
)
print(p1+plot_annotation(title = sample_name))
}
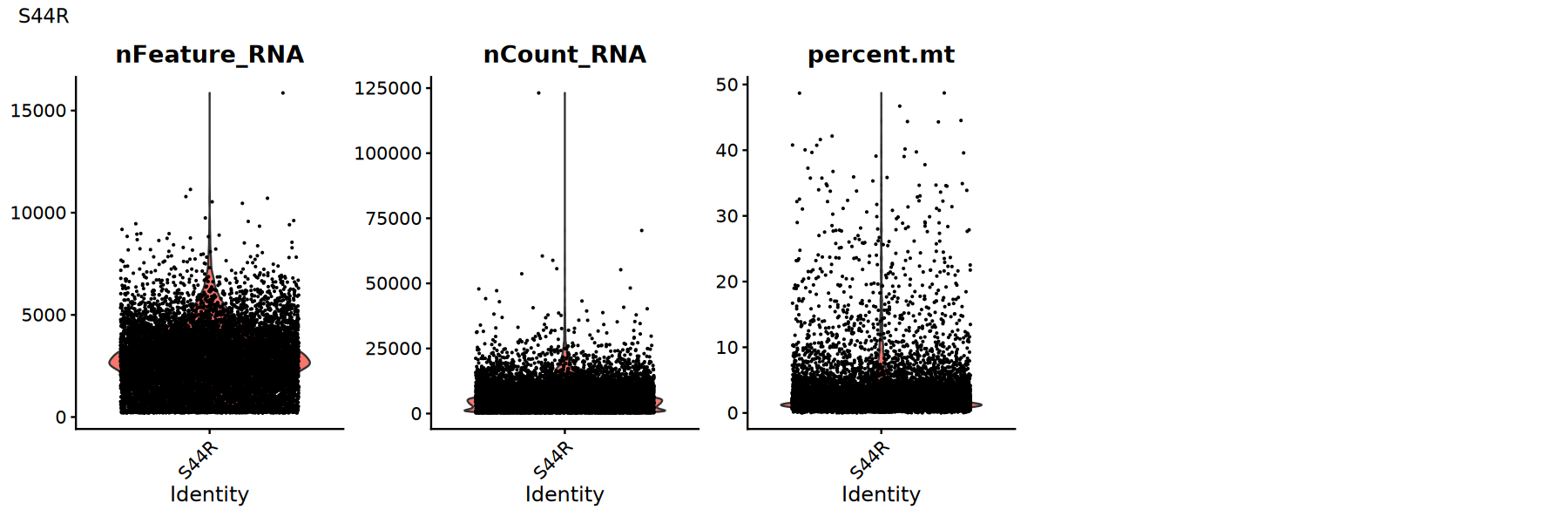
低质量细胞过滤
根据质控指标过滤低质量细胞。具体阈值应根据数据特征进行调整。具体过滤阈值参考上面的小提琴图分布情况。
# 使用lapply进行质量控制过滤
seurat_list <- lapply(seurat_list, function(x) {
# 记录过滤前的细胞数量
cells_before <- ncol(x)
# 质量控制过滤
x <- subset( x,subset = nFeature_RNA > 200 & # 基因数量大于200
nFeature_RNA < 8000 & # 基因数量小于5000
nCount_RNA > 500 & # UMI数量大于500
nCount_RNA < 30000 & # UMI数量小于30000
percent.mt < 20 # 线粒体基因比例小于20%
)
# 记录过滤后的细胞数量
cells_after <- ncol(x)
# 输出过滤信息
cat('过滤完成: 过滤前', cells_before, '个细胞,过滤后', cells_after, '个细胞\n')
return(x)
})过滤完成: 过滤前 11830 个细胞,过滤后 11122 个细胞
多样本合并
在 scRNA-seq 数据分析流程中,多样本合并是整合分析的重要前置步骤。
合并操作的目的:
- 将多个样本数据整合到同一个 Seurat 对象中
- 为后续的批次矫正和整合分析做准备
- 便于与批次矫正后的结果进行对比分析
重要说明:
- 此步骤仅进行简单的数据合并,尚未进行批次效应矫正
- 合并后的对象包含所有样本的原始数据
- 将合并的数据进行标准化、特征选择、PCA 降维和 UMAP 可视化等预处理步骤
# 合并所有样本
suppressWarnings({
suppressMessages({
obj_merge <- merge(seurat_list[[1]], seurat_list[-1], merge.data = FALSE)
obj_merge <- NormalizeData(obj_merge)
obj_merge <- FindVariableFeatures(obj_merge, nfeatures = 2000)
obj_merge <- ScaleData(obj_merge)
obj_merge <- RunPCA(obj_merge)
})
})数据整合
现在我们将多个样本的 scATAC-seq 数据进行整合,并对整合的数据进行降维聚类。scATAC-seq 数据整合主要有两种常用方法:
整合方法选择
CCA(Canonical Correlation Analysis)整合:
- 基于典型相关分析的整合方法
- 通过寻找样本间的共同变异模式进行整合
- 适用于样本间差异较大的情况
- Seurat 包的经典整合方法
Harmony 整合:
- 基于迭代聚类的快速整合方法
- 直接在降维空间中校正批次效应
- 计算效率高,适用于大规模数据
- 保留更多生物学变异信息
方法选择建议
- Harmony:同平台不同样本 scATAC-seq 数据;样本数量较多或数据量大,Harmony 方法更快。
- CCA:运行时间较久,通常批次效应严重的情况下推荐选 CCA,如跨平台的 scATAC-seq 数据
harmony 方法进行整合
注意:harmony 和 CCA 选择其中一种方法进行批次矫正
# 定义harmony整合函数
integrate_harmony <- function(obj) {
DefaultAssay(obj) <- "RNA"
obj <- RunHarmony(obj, "Sample")
obj <- RunUMAP(obj, reduction = "harmony",
dims = 1:30)
#obj <- RunTSNE(obj, reduction = "harmony",dims = 1:30,check_duplicates = FALSE)
# RNA数据聚类
obj <- FindNeighbors(object = obj, reduction = 'harmony', dims = 1:30)
obj <- FindClusters(object = obj, verbose = FALSE, algorithm = 3,resolution = 0.5)
return(obj)
}
#执行harmony整合分析
suppressWarnings({
suppressMessages({
obj_integrated = integrate_harmony(obj_merge)
})
})CCA 方法进行整合
注意:harmony 和 CCA 选择其中一种方法进行批次矫正
# 切换到RNA assay
suppressWarnings({
suppressMessages({
objs <- lapply(seurat_list, function(x) {
DefaultAssay(x) <- "RNA"
return(x)
})
cat('合并后细胞总数:', ncol(obj_merge), '\n')
# 定义CCA整合函数
integrate_cca <- function(objs) {
objs <- lapply(objs, function(x) {
x <- NormalizeData(x, verbose = FALSE)
x <- FindVariableFeatures(x,nfeatures = 2000,selection.method = "vst")
return(x)
})
features <- Seurat::SelectIntegrationFeatures(object.list = objs)
anchors <- Seurat::FindIntegrationAnchors(
object.list = objs,
anchor.features = features
)
obj <- Seurat::IntegrateData(anchorset = anchors)
DefaultAssay(obj) <- "integrated"
obj <- Seurat::ScaleData(obj, verbose = FALSE) %>%
RunPCA(verbose = FALSE)
#对整合后的数据进行UMAP降维
obj <- RunUMAP(obj, reduction = "pca", dims = 1:30)
# RNA数据聚类
obj <- FindNeighbors(object = obj, dims = 1:30)
obj <- FindClusters(object = obj, verbose = FALSE, algorithm = 3,resolution = 0.5)
#obj <- RunTSNE(obj, reduction = "pca", dims = 1:30, check_duplicates = FALSE)
return(obj)
}
# 执行CCA整合
obj_integrated <- integrate_cca(seurat_list)
})
})整合效果评估
比较整合前后的结果,评估整合效果。
评估指标:
- 样本混合程度:不同样本的细胞在 UMAP 图中的分布
- 批次效应去除:技术重复样本的聚集情况
- 生物学信号保留:已知细胞类型的分离度
cat('开始可视化整合结果...', Sys.time(), '\n')
# 对比整合前后效果
suppressWarnings({
suppressMessages({
obj_merge <- RunUMAP(obj_merge, reduction = "pca", dims = 1:30)
})
})
p1 <- DimPlot(obj_merge, group.by = "Sample") +
ggtitle("Before integrate")
p2 <- DimPlot(obj_integrated, group.by = "Sample") +
ggtitle("After integrate")
# 保存对比图
pdf("integration_comparison.pdf", width = 16, height = 12)
print(p1 + p2)
dev.off()
options(repr.plot.width = 20, repr.plot.height = 8)
print(p1 + p2)pdf: 2
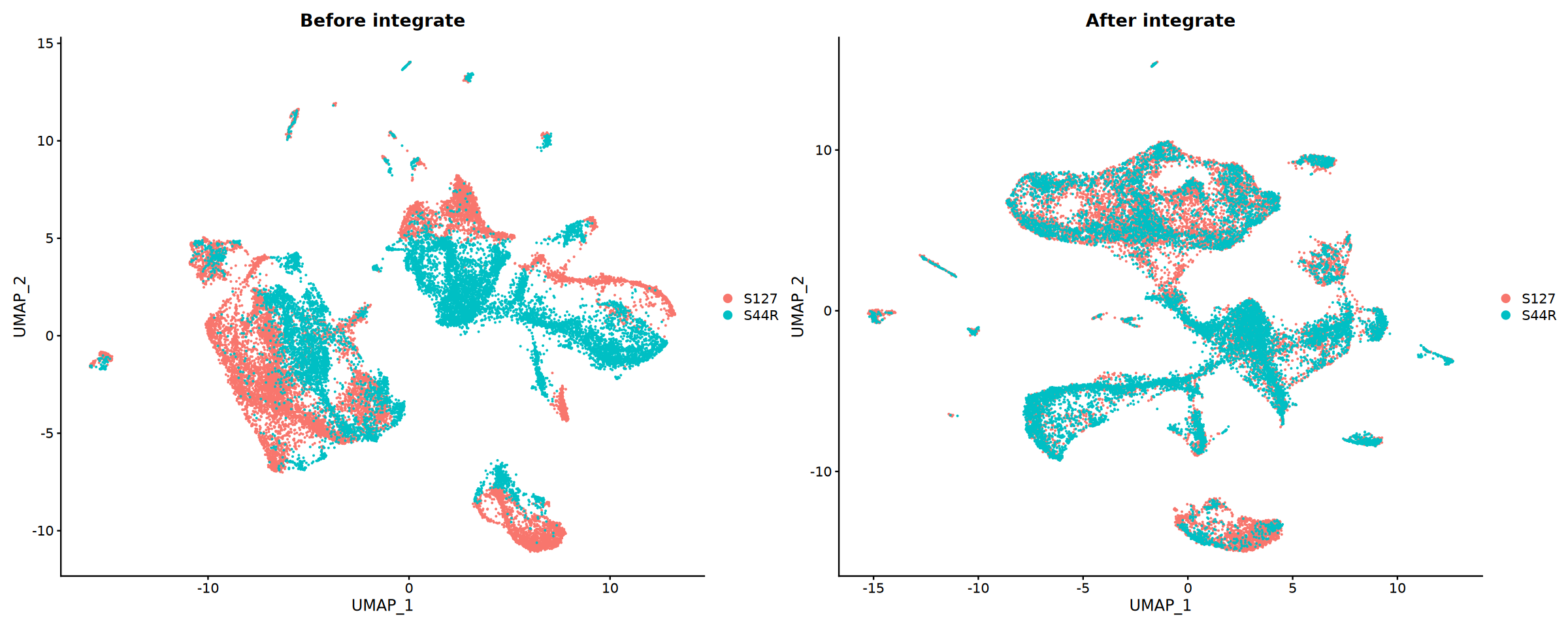
细胞类型注释
基于 marker 基因表达进行细胞类型注释。先对各聚类做差异表达以获取候选 marker,再结合文献与数据库进行判读。
定义 marker 基因
整理常见细胞类型的参考 marker 集合(来源可包括文献、PanglaoDB、CellMarker 等),用于后续可视化与判读。
eye_marker_integrated <- list(
# ========== 光感受器细胞 ==========
"Rod_Photoreceptors" = c("RHO", "PDE6B", "CNGB1", "PDE6A", "NR2E3", "REEP6",
"RCVRN", "SAG", "NEB", "SLC24A3", "TRPM1"),
"Cone_Photoreceptors" = c("ARR3", "GNAT2", "OPN1SW", "TRPM3"),
# ========== 视网膜神经元 ==========
"Bipolar_Cells" = c("VSX1", "OTX2", "GRM6", "PRKCA",
"DLG2", "NRXN3", "RBFOX3", "FSTL4"),
"Amacrine_Cells" = c("GAD1", "GAD2", "C1QL2", "TFAP2B", "SLC32A1"),
"Horizontal_Cells" = c("ONECUT1", "LHX1", "CALB1", "NFIA"),
"Retinal_Ganglion_Cells" = c("RBPMS", "THY1", "NEFL", "POU4F2"),
# ========== 胶质细胞 ==========
"Muller_Glia" = c("SLC1A3", "RLBP1", "SOX9", "CRYAB"),
"Astrocytes" = c("GFAP", "AQP4", "S100B"),
"Microglia" = c("TMEM119", "C1QA", "AIF1", "CX3CR1", "CD74",
"PTPRC", "CSF1R", "CCL3L1", "SPP1", "P2RY12"),
# ========== 血管与支持细胞 ==========
"Endothelial" = c("FLT1", "PODXL", "PLVAP", "KDR", "EGFL7",
"CLDN5", "PECAM1", "CDH5"),
"Pericytes" = c("PDGFRB", "RGS5", "CSPG4", "STAB1"),
# ========== 上皮细胞 ==========
"RPE" = c("RPE65", "BEST1", "PMEL", "TYRP1", "DCT", "SLC38A11"),
#"Corneal_Epithelial" = c("KRT12", "KRT3"),
"Corneal_Endothelial" = c("SLC4A11", "COL8A2", "ATP1A1"),
"Conjunctival_Epithelial" = c("KRT13", "KRT19", "MUC5AC"),
"Retinal_Progenitors" = c("VSX2", "PAX6","SOX2", "HES1", "NOTCH1"), # 神经视网膜增殖期
# ========== 晶状体细胞 ==========
"Lens_Epithelial" = c("CRYAA", "BFSP1", "MIP"),
"Lens_Fiber" = c("CRYGS", "LIM2", "FN1"),
# ========== 其他细胞 ==========
"Melanocytes" = c("TYR", "MLANA", "MITF"),
#"Erythrocytes" = c("HBB", "HBA1", "HBA2"),
"ECM" = c("COL1A1", "COL3A1", "COL12A1", "COL1A2", "COL6A3")#,
#"Others" = c("TTN", "CLCN5", "DCC", "MIAT")
)Marker 基因表达可视化
在不同聚类中展示候选 marker 的表达模式(小提琴图/点图/FeaturePlot),辅助确认类型归属。
cat('开始marker基因表达可视化...', Sys.time(), '\n')
# 创建DotPlot展示marker基因表达
DefaultAssay(obj_integrated)="RNA"
p_dot <- DotPlot(
obj_integrated,
features = eye_marker_integrated,
cols = c("#f8f8f8","#ff3472"),
dot.scale = 8
) +
RotatedAxis() +
ggtitle("Marker genes expression") +
theme(axis.text.x = element_text(angle = 45, hjust = 1))
# 保存DotPlot
pdf("marker_genes_dotplot.pdf", width = 16, height = 8)
print(p_dot)
dev.off()
options(repr.plot.width = 26, repr.plot.height = 8)
print(p_dot)
cat('marker基因表达可视化完成!', Sys.time(), '\n')pdf: 2
marker基因表达可视化完成! 1755238807
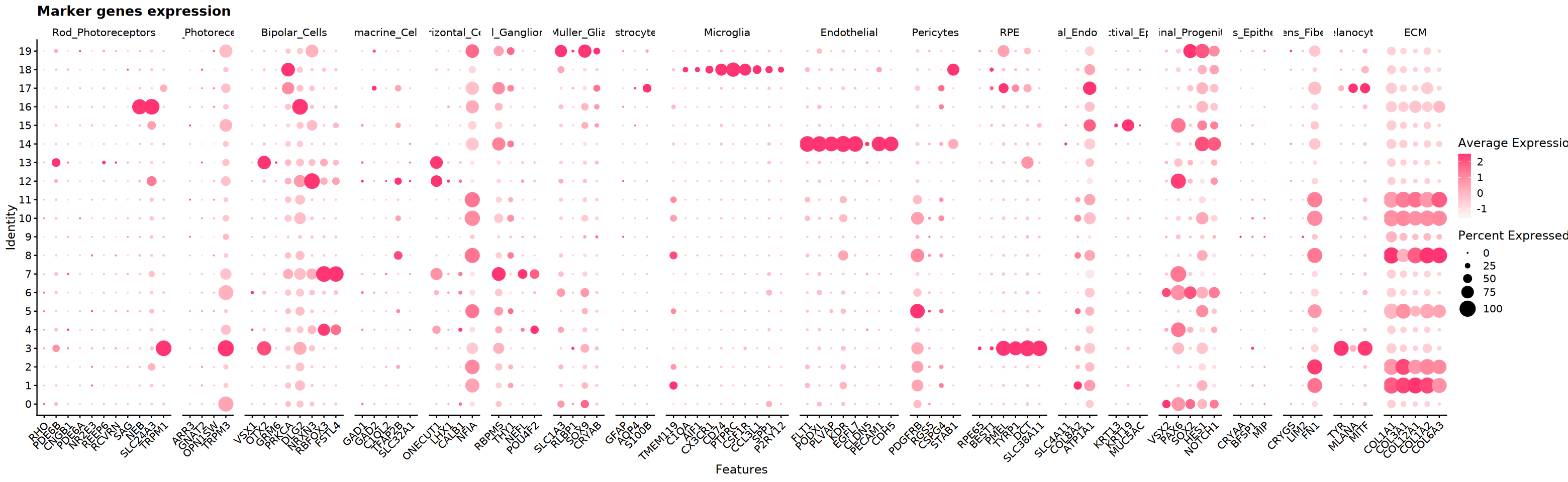
p3 <- DimPlot(obj_integrated,group.by = "seurat_clusters",label = T, cols = my36colors)+ggtitle("Clusters")
options(repr.plot.width = 10, repr.plot.height = 8)
print(p3)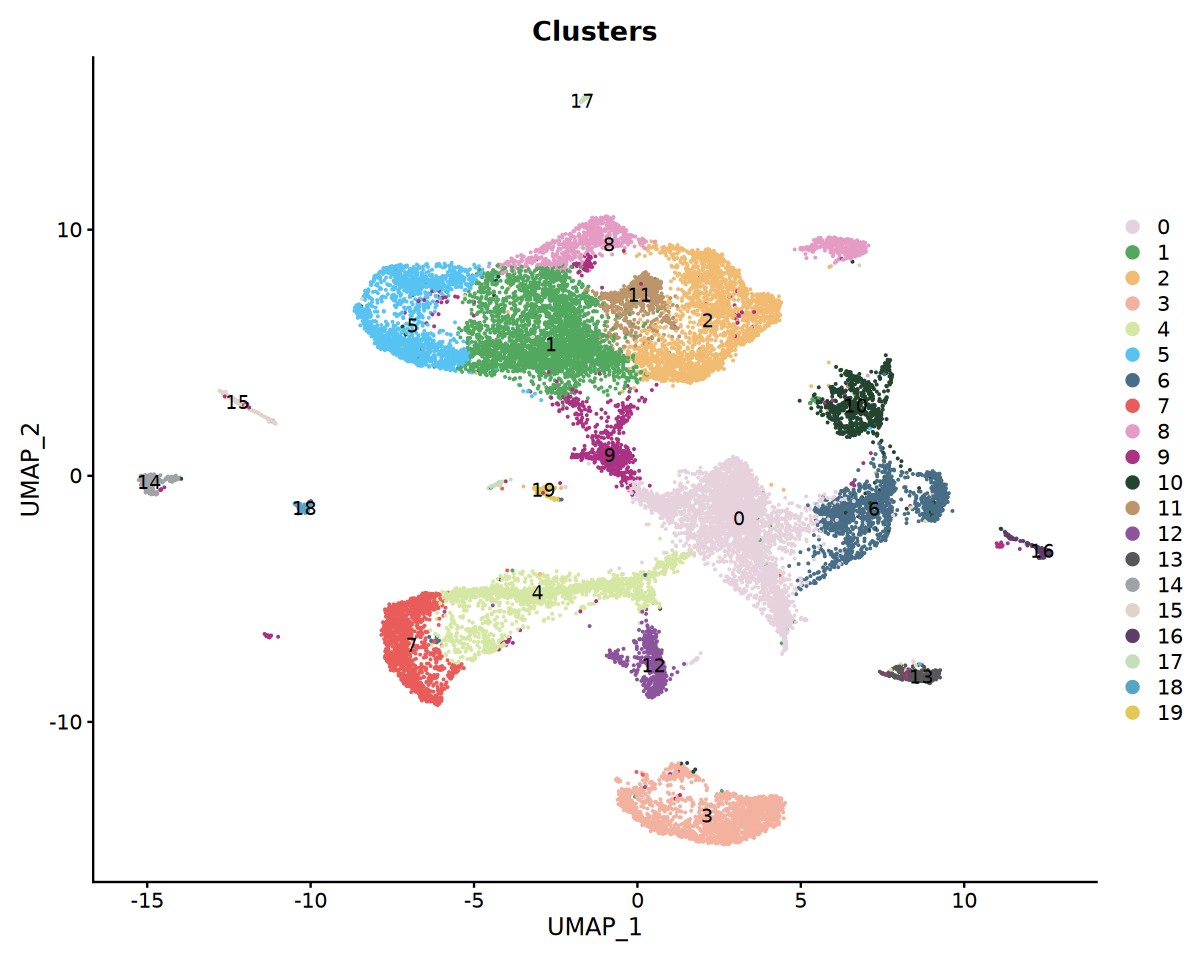
细胞类型注释
结合聚类层面差异基因与参考 marker,综合注释细胞类型;必要时细化到亚型或回退调整分辨率。
cat('开始细胞类型注释...', Sys.time(), '\n')
# 基于聚类结果进行细胞类型注释(需要根据实际的marker基因表达情况调整)
# 这里提供一个示例,实际使用时需要根据DotPlot结果进行调整
celltype_mapping <- c(
"0" = "Retinal_Progenitors",
"1" = "ECM",
"2" = "ECM",
"3" = "PRE",
"4" = "Bipolar_Cells",
"5" = "ECM",
"6" = "Retinal_Progenitors",
"7" = "Bipolar_Cells",
"8" = "ECM",
"9" = "Conjunctival_Epithelial",
"10" = "ECM",
"11" = "ECM",
"12" = "Bipolar_Cells",
"13" = "Doublets",
"14" = "Endothelial",
"15" = "Conjunctival_Epithelial",
"16" = "Rod_Photoreceptors",
"17" = "Astrocytes",
"18" = "Microglia",
"19" = "Muller_Glia"
)
# 应用细胞类型注释
obj_integrated$celltype <- recode(
obj_integrated$seurat_clusters,
!!!celltype_mapping
)细胞类型注释可视化
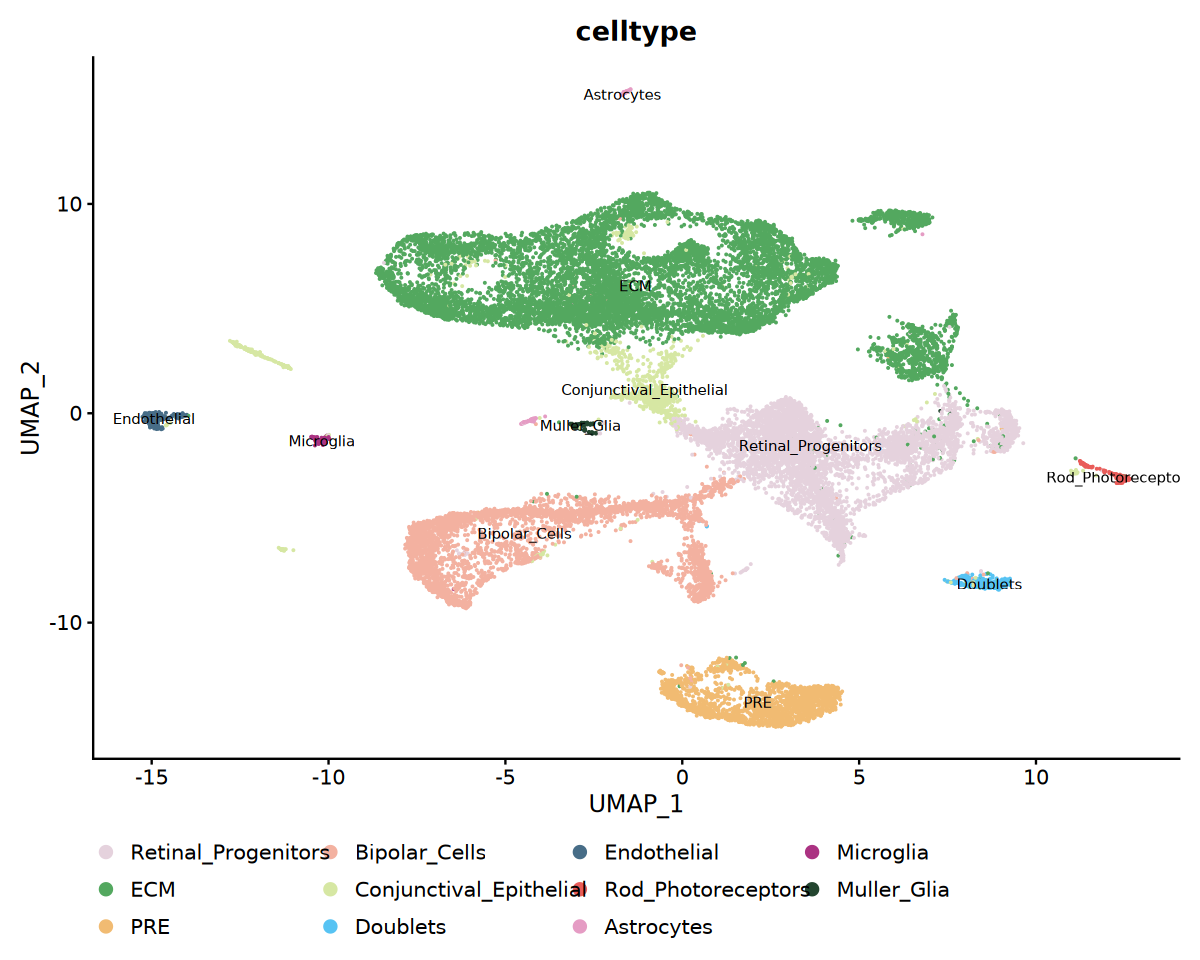
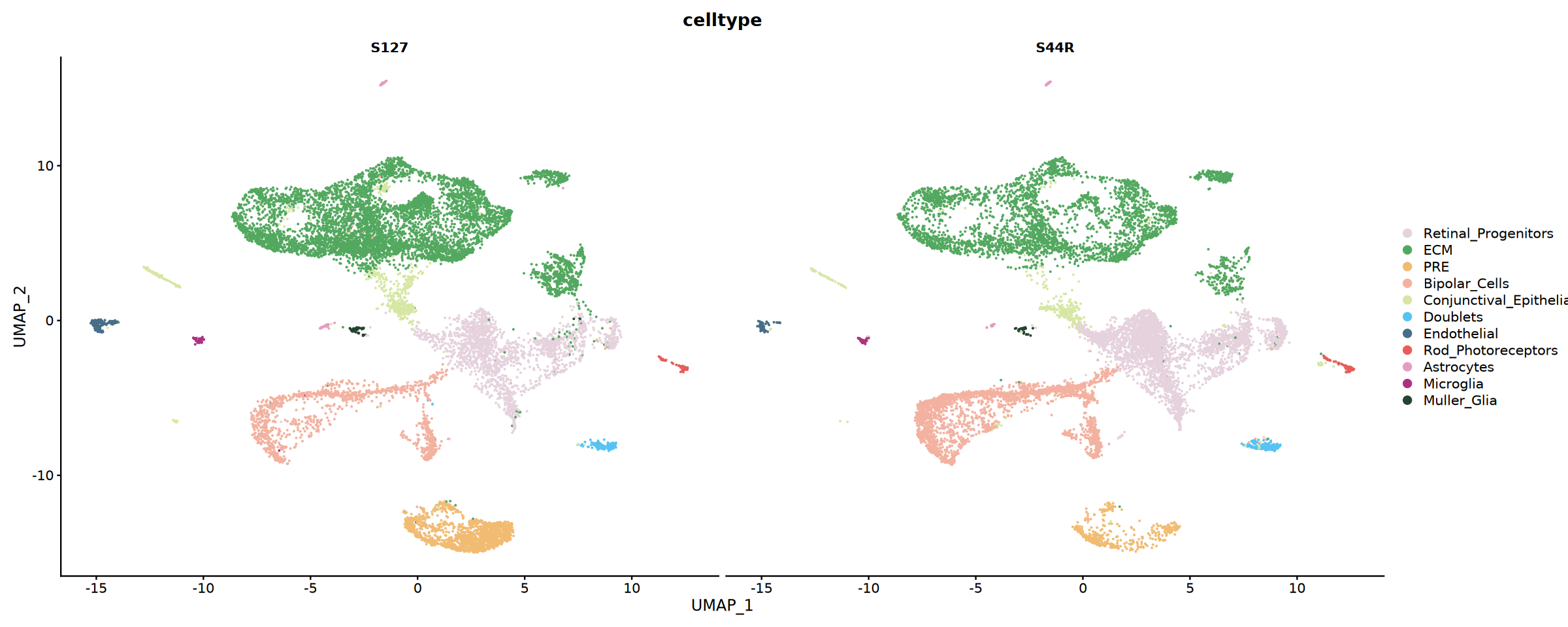
以 UMAP 着色或按样本分面展示注释结果,并可统计各细胞类型的样本比例用于对比分析。
options(repr.plot.width = 10, repr.plot.height = 8)
# 细胞类型UMAP可视化
p1 <- DimPlot(
obj_integrated,
reduction = "umap",
group.by = "celltype",
label = TRUE,
label.size = 3,
cols = my36colors
) +
theme(legend.position = "bottom")
# 按样本分组展示细胞类型分布
p2 <- DimPlot(
obj_integrated,
reduction = "umap",
group.by = "celltype",
split.by = "Sample",
cols = my36colors,
ncol = 2
)
# 保存细胞类型注释图
pdf("celltype_annotation.pdf", width = 16, height = 12)
print(p1)
print(p2)
dev.off()
print(p1)
options(repr.plot.width = 20, repr.plot.height = 8)
print(p2)pdf: 2
结果保存
保存整合后的对象与关键图表,建议同时保存中间对象以便复现与后续复查。
# 保存整合后的Seurat对象
saveRDS(obj_integrated, file = "scRNA_scRNA_integrated.rds")总结
本教程演示了 scRNA-seq 多样本整合的完整流程,涵盖:
- 数据读取:导入 10x 格式表达矩阵,创建 Seurat 对象
- 质量控制:计算质控指标并筛除低质量细胞
- 数据预处理:归一化与高变基因识别
- 多样本合并:汇总样本并保留来源信息
- 数据整合:使用 CCA/RPCA 消除批次效应
- 降维与聚类:PCA、UMAP 及图聚类
- 细胞类型注释:基于 marker 的判读与可视化
最佳实践:
- 动态调整质控与整合参数(如阈值、dims、锚点数)
- 结合生物学先验与公共数据库进行注释
- 按阶段保存对象,确保分析可追溯与复现
后续分析建议:
- 组织类型偏好性分析
- 差异富集分析
- 拟时序分析
- 细胞互作分析
sessionInfo()Platform: x86_64-conda-linux-gnu (64-bit)
Running under: Debian GNU/Linux 12 (bookworm)
Matrix products: default
BLAS/LAPACK: /jp_envs/envs/common/lib/libopenblasp-r0.3.29.so; LAPACK version 3.12.0
locale:
[1] LC_CTYPE=C.UTF-8 LC_NUMERIC=C LC_TIME=C.UTF-8
[4] LC_COLLATE=C.UTF-8 LC_MONETARY=C.UTF-8 LC_MESSAGES=C.UTF-8
[7] LC_PAPER=C.UTF-8 LC_NAME=C LC_ADDRESS=C
[10] LC_TELEPHONE=C LC_MEASUREMENT=C.UTF-8 LC_IDENTIFICATION=C
time zone: Asia/Shanghai
tzcode source: system (glibc)
attached base packages:
[1] stats graphics grDevices utils datasets methods base
other attached packages:
[1] future_1.40.0 harmony_1.2.3 Rcpp_1.0.14 patchwork_1.3.0
[5] ggplot2_3.5.2 dplyr_1.1.4 SeuratObject_4.1.4 Seurat_4.4.0
[9] repr_1.1.7
loaded via a namespace (and not attached):
[1] deldir_2.0-4 pbapply_1.7-2 gridExtra_2.3
[4] rlang_1.1.5 magrittr_2.0.3 RcppAnnoy_0.0.22
[7] spatstat.geom_3.3-6 matrixStats_1.5.0 ggridges_0.5.6
[10] compiler_4.3.3 png_0.1-8 vctrs_0.6.5
[13] reshape2_1.4.4 stringr_1.5.1 pkgconfig_2.0.3
[16] crayon_1.5.3 fastmap_1.2.0 labeling_0.4.3
[19] promises_1.3.2 ggbeeswarm_0.7.2 purrr_1.0.4
[22] jsonlite_2.0.0 goftest_1.2-3 later_1.4.2
[25] uuid_1.2-1 spatstat.utils_3.1-3 irlba_2.3.5.1
[28] parallel_4.3.3 cluster_2.1.8.1 R6_2.6.1
[31] ica_1.0-3 stringi_1.8.7 RColorBrewer_1.1-3
[34] spatstat.data_3.1-6 reticulate_1.42.0 parallelly_1.43.0
[37] spatstat.univar_3.1-2 lmtest_0.9-40 scattermore_1.2
[40] IRkernel_1.3.2 tensor_1.5 future.apply_1.11.3
[43] zoo_1.8-14 R.utils_2.13.0 base64enc_0.1-3
[46] sctransform_0.4.1 httpuv_1.6.15 Matrix_1.6-5
[49] splines_4.3.3 igraph_2.0.3 tidyselect_1.2.1
[52] abind_1.4-5 spatstat.random_3.3-3 codetools_0.2-20
[55] miniUI_0.1.1.1 spatstat.explore_3.4-2 listenv_0.9.1
[58] lattice_0.22-7 tibble_3.2.1 plyr_1.8.9
[61] withr_3.0.2 shiny_1.10.0 ROCR_1.0-11
[64] ggrastr_1.0.2 evaluate_1.0.3 Rtsne_0.17
[67] survival_3.8-3 polyclip_1.10-7 fitdistrplus_1.2-2
[70] pillar_1.10.2 KernSmooth_2.23-26 plotly_4.10.4
[73] generics_0.1.3 sp_2.2-0 IRdisplay_1.1
[76] munsell_0.5.1 scales_1.3.0 globals_0.16.3
[79] xtable_1.8-4 glue_1.8.0 lazyeval_0.2.2
[82] tools_4.3.3 data.table_1.17.0 pbdZMQ_0.3-13
[85] RANN_2.6.2 leiden_0.4.3.1 Cairo_1.6-2
[88] cowplot_1.1.3 grid_4.3.3 tidyr_1.3.1
[91] colorspace_2.1-1 nlme_3.1-168 beeswarm_0.4.0
[94] vipor_0.4.7 cli_3.6.4 spatstat.sparse_3.1-0
[97] viridisLite_0.4.2 uwot_0.2.3 gtable_0.3.6
[100] R.methodsS3_1.8.2 digest_0.6.37 progressr_0.15.1
[103] ggrepel_0.9.6 htmlwidgets_1.6.4 farver_2.1.2
[106] R.oo_1.27.0 htmltools_0.5.8.1 lifecycle_1.0.4
[109] httr_1.4.7 mime_0.13 MASS_7.3-60.0.1